





서 론
Gamma-aminobutyric acid(GABA)는 단백질을 구성하지 않는 기능성 아미노산이며, glutamate decarboxylase(GAD)에 의해 L-glutamic acid로부터 생산된다(Kook and Cho, 2013). GABA는 이뇨제, 혈압 강하제, 진정제, 항당뇨 효과 등의 여러 생리학적 기능을 가지며, 특히, 뇌 세포에서 억제성 신경전달물질로 이용되어 산소 공급을 증가하고 뇌 혈류를 활성화함으로써 뇌 세포 대사를 촉진한다(Cho et al., 2007; Kook and Cho, 2013; Lee et al., 2018). GABA는 성장 초기에 뇌의 해마와 피질에 주요한 자극을 주기 때문에 자극성 신경전달물질로도 활동하며, 뇌가 성장하면서 흥분을 조절하는 억제성 신경전달물질로 활동하게 된다(Li and Xu, 2008). 유산균은 GABA를 생산하는 주요 생물체 중 하나이며, 이는 Lactobacillus brevis와 같은 Lactobacillus 속의 여러 유산균이 glutamate decarboxylase 시스템을 가지고 있기 때문이다(Lee et al., 2018; Lim et al., 2018; Sa et al., 2015; Wu and Shah, 2017). GABA 생산 유산균은 여러 발효 식품으로부터 분리 가능하며, GABA를 생산하는 유산균을 이용한 기능성 발효식품 개발 연구가 폭넓게 진행되고 있다. 예를 들면, 묵은지 김치로부터 GABA 생산능이 높은 유산균을 분리하여 신 김치 발효에 이용한다거나, GABA 생산 능력을 가지는 유산균을 유제품 발효의 스타터로 이용하여 GABA가 첨가된 기능성 식품을 생산하는 것이다(Cho et al., 2011; Lee et al., 2018; Wu and Shah, 2017). 최근에는 유산균의 섭취가 항 우울증, 항 불안증 효과와 스트레스 반응 해소 효과가 있다는 동물실험과 임상연구도 보고되고 있으며, GABA와의 연관성을 언급하여 유산균과 뇌, GABA 사이에 상관관계에 대한 연구의 폭이 더 커질 것으로 예상된다(Barros-Santos et al., 2020). Cho 등(2007)의 연구에서는 김치로부터 분리한 GABA 생산 유산균의 배양 추출물이 신경독소 유도성 세포 사멸에 대하여 신경 세포 보호 효과가 있다는 결과를 보고하였다. 또한, 장내 미생물과 뇌 기능의 연관성은 오래 전부터 관심 있게 연구되어 왔으며, Naseribafrouei 등(2014)은 인간 분변 미생물 군집과 우울증과의 상관관계에 대한 임상연구를 진행하여, 우울증 환자의 분변에서 낮은 발현을 보여준 Oscillibacter 속의 최종 대사물질인 발레르산이 신경전달물질인 GABA와 유사하다고 보고하였으며, 우울증과 유의적 상관관계가 있을 것으로 추측하였다. 유산균 섭취와 뇌 건강 사이의 연구는 특히, Lactobacillus rhamnosus JB-1(이하 JB-1)섭취에 따른 장관과 뇌, 행동 변화에 대한 연구가 많이 보고되고 있다. Bravo 등(2011)은 JB-1의 불안 해소 효과가 GABA에 의한 메커니즘과 연관되어 있다고 추측하고 있으며, 중심 GABA 수용체 발현의 변화가 불안증과 우울증의 발병 원인이며, 만성적 JB-1 치료가 GABA 수용체 발현의 변화를 유도하여 스트레스 유도 물질인 코르티코스테론 수준과 불안증, 우울증 관련 행동을 감소하였다고 보고하였다. Bharwani 등(2017)은 JB-1의 경구 섭취에 따른 마우스의 장내 미생물 변화와 함께 면역 반응 변화, 스트레스 변화 등을 연구하여 JB-1의 스트레스 보호 효과에 대한 연구 결과도 보고하였다. Janik 등(2016)은 마우스에서 JB-1의 치료 동안에 GABA, 글루탐산, 글루타민 등의 신경전달물질의 증가와 치료 중단 후에도 글루탐산과 글루타민은 계속 증가한다는 결과를 통해 불안증, 우울증과 관련된 특정 뇌 구역의 활성과 기능에 유산균이 관여한다는 것을 보고하였고, Kochalska 등(2020)은 스트레스를 받은 렛의 뇌 반응 관찰을 통해 GABA, 글루탐산, 글루타민 등의 신경전달물질의 변화가 JB-1에 의해 스트레스 받기 이전처럼 호전된다는 연구 결과를 발표하였다. 따라서 본 연구는 GABA를 생산하는 유산균의 섭취가 장내 미생물과 대사, 뇌 기능과 행동에 직접적으로 또는 간접적으로 어떠한 영향을 줄 수 있을지에 대한 의문에 답을 찾기 위한 출발선으로 전통발효식품인 김치로부터 GABA 생산 유산균을 분리하여 그 특성을 알아보고자 한다.
재료 및 방법
일반가정집에서 담근 배추김치 7종, 물김치 2종, 열무김치, 갓김치, 갈치김치 각 1종, 경북 경산시 하양읍 식당 6곳에서 얻은 배추김치 6종, 식당 3곳에서 얻은 물김치, 동치미, 깍두기 각 1종 등 총 21종의 김치로부터 유산균을 분리하였다. 김치국물을 0.9% NaCl로 단계 희석하여 BCP 평판배지(0.5% glucose, 2% tryptone, 0.5% yeast extract, 0.4% NaCl, 0.15% sodium acetate, 40 mg/L bromocresol purple)에 도말한 후, 30℃에서 24시간 동안 배양하였다. 배양 후, 노란색 환을 나타내는 콜로니를 무작위로 추출하여 순수 분리하고, glycerol stock을 만들어 –70℃에서 보관하면서 실험에 이용하였다.
김치로부터 분리한 유산균들은 MRS 액체배지에서 30℃, 24시간 배양한 후, 원심분리와 0.20 μm syringe filter를 이용하여 균체를 제거한 배양상등액을 얻었다. 배양상등액은 Tsukatani 등(2005)의 microtiter plate assay를 참고하여 GABA 농도를 측정하였다. 먼저, 80 mM Tris-HCl buffer(pH 9.0)에 750 mM sodium sulfate, 10 mM dithiothreitol, 1.4 mM NADP+, 2 mM α-ketoglutarate, 30 μg GABase를 포함하여 microtiter plate에 90 μL씩 분주하였다. 다음은 앞서 준비한 배양상등액을 10 μL씩 넣고 30℃에서 60분 동안 반응시킨 후, 340 nm에서 흡광도를 측정하였다. 표준물질 GABA를 5.0×10-4~1.0×10-2 M 범위에서 같은 방법으로 흡광도를 측정하여 얻은 표준곡선을 이용해 GABA 농도를 계산하였다. 표준물질 blank는 증류수, 배양상등액 blank는 MRS 배지를 GABase 없이 측정하였다. GABA 농도에 따라 1차 선별한 유산균은 MRS에서 48시간, 5% monosodium glutamic acid(MSG)를 첨가한 MRS에서 24, 48시간 배양한 후, 배양상등액을 얻어 같은 방법으로 GABA 농도를 측정하여 GABA 농도가 가장 높은 균주를 최종 선발하였다.
결과 및 고찰
김치로부터 703개의 균주를 분리하였다. 703개의 균주를 MRS 배지에 배양하여 상등액을 얻었고, GABA 농도를 측정하여 표준곡선 범위 안에 드는 균주 17개를 1차 선별하였다. 1차 선별한 유산균들을 MRS 배지와 5% MSG를 포함하는 MRS 배지에 24, 48시간 배양하여 얻은 상등액으로 2차 실험을 하였고, 모든 시료에서 GABA를 가장 높게 생산하는 두 개의 균주를 최종 선발하였다. 두 균주는 K991과 K998이며, K991은 MRS 배지에서 24, 48시간 배양했을 때, 각각 3.52 mM, 4.24 mM 농도로 측정되었고, K998은 1.66 mM, 3.85 mM로 측정되었다. 5% MSG가 포함된 MRS 배지에서는 24, 48시간 배양했을 때, K991의 경우 각각 2.43 mM, 9.60 mM, K998의 경우 각각 2.68 mM, 9.00 mM로 측정되었다(Table 1).
16S rRNA 유전자 염기서열 분석을 통해 K991, K998은 각각 99.93%, 99.87% 일치율을 보여준 Lactococcus lactis로 동정되었으며, Lactococcus lactis K991, Lactococcus lactis K998로 명명하였다. Table 2와 Table 3에서 보는 바와 같이 K991과 K998은 공통적으로 Lactococcus lactis subsp. lactis bv. diacetylactis strain SD96과 Lactococcus lactis strain IL1403(이하 IL1403)에 대해 99% 이상의 일치율을 보였으며, 두 균주는 모두 치즈 생산에 스타터로 이용되었다(Dorau et al., 2020; Bolotin et al., 2001). 그 외에 Lactococcus lactis strain IL6288의 경우, IL1403에서 프로파지가 제거된 균주로 유제품 스타터로서 가능성을 보고하였다(Bolotin et al., 2019). Table 2와 Table 3에 표시된 것 외에 NCBI BLAST 검색을 통해 K991과 K998에 가장 높은 일치율(각 99.93%, 99.87%)을 보이는 Lactococcus lactis 균주 목록 중 공통적인 균주들이 많았으며, 그 중에서 Lactococcus lactis subsp. lactis NCDO 2118은 냉동 콩에서 분리한 유산균으로 GABA를 생산하는 것으로 보고되었다(Oliveira et al., 2014). K991 BLAST 검색 결과에는 없었던 Lactococcus lactis strain CJNU 3001(이하 CJNU 3001)은 K998과 99.87%의 일치율을 보여주었고, CJNU 3001은 항 Staphylococcus aureus 박테리오신을 생산하는 균주로 보고되었으며(Yang and Moon, 2018), 김치에서 분리되었다는 점에서 K998과 접점을 보여주었다.
| Strain | Identity (%)1) |
|---|---|
| Lactococcus lactis subsp. lactis bv. diacetylactis strain SD96 | 99.93 |
| Lactococcus lactis strain IL1403 | 99.93 |
| Lactococcus lactis strain IL6288 | 99.93 |
| Strain | Identity (%)1) |
|---|---|
| Lactococcus lactis strain CJNU 3001 | 99.87 |
| Lactococcus lactis subsp. lactis bv. diacetylactis strain SD96 | 99.87 |
| Lactococcus lactis strain IL1403 | 99.87 |
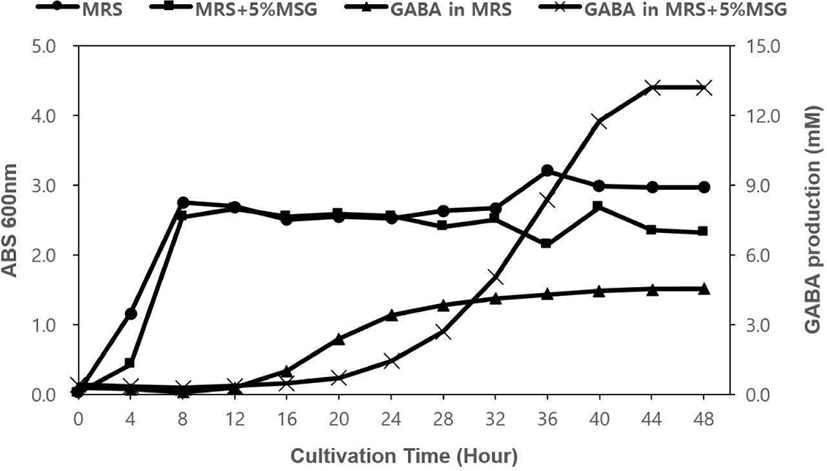
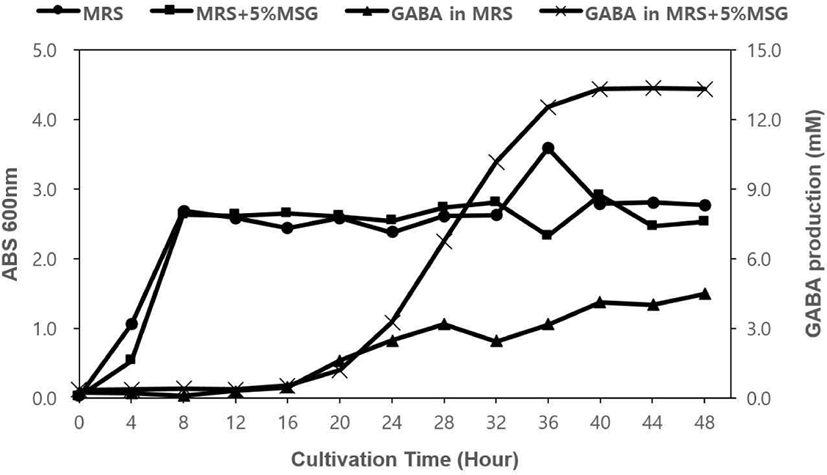
L. lactis K991과 L. lactis K998은 MRS 액체 배지와 5% MSG를 포함한 MRS 액체 배지에서 30℃, 48시간 동안 배양하면서 생육에 따른 GABA 생산 능력을 측정하였다. L. lactis K991의 경우, MRS와 5% MSG를 포함하는 MRS에서 비슷한 경향으로 성장하였다. 배양 4시간 이후에 대수기에 접어들어 8시간 이후 정지기에 도달하였다. 균주가 성장하는 경향은 비슷했지만, GABA를 생산하는 능력에는 배지에 따라 차이가 있었다. MRS에서 배양했을 때는 배양 12시간 이후부터 GABA 생산량이 서서히 늘어나서 28시간 이후부터 생산 증가량이 완만하게 유지되었다. 반면, 5% MSG가 포함된 MRS에서는 배양 16시간 이후부터 생산량이 늘어나기 시작하여 28시간 이후부터 급격하게 증가하였고, 44시간에 최고치에 도달하였다. 이 때, GABA 생산량이 13.20 mM로 MRS에서 같은 시간에 생산된 양인 4.53 mM보다 약 3배나 높은 생산력을 보여주었다(Figure 1). L. lactis K998의 경우, L. lactis K991의 성장과 비슷하게 두 가지 배지에서 모두 배양 4시간 이후부터 대수기가 시작되었고, 8시간 이후에 정지기로 접어들었다. MRS에서 GABA 생산은 16시간 이후부터 서서히 증가하였고, 5% MSG를 포함한 MRS에서도 역시 16시간 이후부터 GABA 생산이 증가하기 시작하여 20시간 이후부터 급격하게 증가하였고, 40시간에 최대치에 도달하여 유지하였다. L. lactis K991과 마찬가지로 두 가지 배지 사이의 GABA 최대 생산량이 MRS에서 4.49 mM, 5% MSG를 포함한 MRS에서 13.35 mM로 약 3배 차이가 났다(Figure 2). 종합적으로 L. lactis K991과 L. lactis K998의 GABA 생산은 정지기에 접어들어 증가하기 시작하였고, MSG를 포함하였을 때 생산량이 높게 나타났다. Cho 등(2011), Cho 등(2007), Lim 등(2017)의 결과와 같이 대수 증식이 시작되면서 GABA 생산도 증가하여, 정지기에 접어들어 최대치의 GABA 생산을 보여주는 것과는 대비되었다. 또한 MRS 배지와 MSG를 포함한 MRS 배지에서 성장의 특징이 유사하지만 GABA 생산은 기질이 풍부한 후자에서 높게 나타난 것으로 보면, 기존 대부분의 GABA 생산 균주 분리가 MSG를 포함하는 배지에서 진행된 이유를 알 수 있었다. 그러나, 유산균을 섭취하였을 때, MSG와 같은 GABA를 생산할 수 있는 기질이 풍부한 환경일지는 알 수 없기에 장내 환경이나 더 극한의 환경에서도 GABA를 생산할 수 있다면 더 가치가 있을 것이며, 그에 대한 연구도 필요할 것으로 사료된다.
요약
본 연구는 GABA 생산 프로바이오틱스 개발을 위해 김치로부터 GABA를 생산하는 유산균을 분리하여 그 특성을 알아보고자 하였다. 먼저, 우리는 21종의 김치로부터 703개의 유산균을 분리하였고 이들 중에서 GABA 생산능력이 높은 유산균 17개를 1차 선별하였다. 이들 중 GABA 생산능력이 우수한 두 균주 K991과 K998을 최종 선발하였다. 16S rRNA 유전자 염기서열 분석 결과, K991과 K998은 각각 99.93%, 99.87%의 일치율을 보여준 Lactococcus lactis로 동정되었다. 30℃에서 MSG를 포함한 MRS 배지에 두 균주를 배양했을 때 12시간 이후에 GABA를 생산하기 시작했으며, 40시간 이후에 최대 생산량을 보여주었다. 반면에 MSG가 첨가되지 않았을 때는 약 3배 정도 낮은 GABA 생산량을 보였다.